Рис.1
| ATHLETE.RU :: Форум :: Книги | Настройка |
Размышляя над названием будущей статьи, я не случайно остановил свой выбор на том варианте, что написан чуть выше - читатель без труда узнает в нем коллаж, составленный из заглавий двух, пожалуй, наиболее популярных в среде спортсменов-любителей, книг о бодибилдинге. "Думай! Бодибилдинг без стероидов" Стюарта МакРоберта и "Супертренинг" Майка Ментцера всколыхнули мир любительского спорта и перевернули, казалось, устоявшиеся представления о теории тренинга. Точнее было бы сказать, что Ментцер впервые попытался создать хоть какую то теорию, до него большинство популярных книг и статей о бодибилдинге были всего лишь сборниками всевозможных и зачастую противоречивых принципов тренировок, и каталогами известных упражнений с тяжестями. Ментцер призвал рассматривать бодибилдинг как науку, но, почему-то, выбрал в качестве основы не физиологию, а философию и логику. Как некогда Евклид создал свою геометрию, опираясь на ряд аксиом о свойствах пространства, так и Ментцер создал свой "Супертренинг" опираясь на аксиому о роли последнего "отказного" повторения в механизме запуска роста мышц, не потрудившись дать никакого физиологического объяснения своей гипотезе. Но, как мы знаем, помимо геометрии Евклида существуют геометрии Лобачевского и Минковского, основанные на других аксиомах, но так же внутренне вполне не противоречивые и логичные. Вдохновленный отменным стилем и непоколебимой уверенностью автора "Супертренинга" в своей правоте, нарастив, следуя его советам, 10 килограмм "натуральных" мышц за полгода, я стал ярым сторонником идей Ментцера. Решив найти физиологическое подтверждение аксиомы "учителя", я с головой окунулся в новую для себя область познания - физиологию и биохимию человека. Результат оказался для меня неожиданным, но об этом чуть позже.
Позволю себе обратить внимание читателей на то чудовищное положение, в котором оказалась теория современного "железного" спорта. Все спортивные журналы полны статей с новыми супермодными системами тренинга. "Движение должно быть мощным и взрывным" - утверждают одни. "Только медленное подконтрольное движение" - противоречат им другие. "Хочешь нарастить массу - работай с большими весами". "Вес снаряда не имеет значения - главное техника и ощущение работы мышцы". Тренироваться шесть раз в неделю утром и вечером советует Арнольд Шварценеггер. Запрещает появляться в зале чаще двух раз в неделю своим ученикам Майк Ментцер. Профи расписывают комплексы из шести упражнений для бицепса. МакРоберт призывает вообще не тренировать руки изолированными упражнениями. Пауэрлифтеры во время своих циклов почти никогда не работают до отказа. Ментцер уверяет, что работа не до отказа - это зря потраченное время. Профи из команды Джо Вейдера советуют идти гораздо дальше отказа с помощью форсированных повторений и "стриптиза". Это перечисление можно продолжить до бесконечности, но поражает не обилие взаимоисключающих принципов тренинга, а то, что у каждого из них находятся свои сторонники, сумевшие получить от их использования результат. Этот факт позволил в широких кругах распространиться мнению, что системы нет. Я же утверждаю, что система есть! И терпеливый читатель вскоре сможет сам убедиться в этом.
Слабым местом любой методики тренинга является отсутствие у ее автора четкого представления о причинах роста мышц. Некоторые вовсе не задумываются над этим вопросом, некоторые ограничиваются абстрактной идеей о тренировочном стрессе. Полагая, что это беда только популярной спортивной литературы я надеялся найти ответ в серьезных научных исследованиях в области физиологии мышечной деятельности, но был разочарован. Точными сведениями о том, что именно происходит в мышце во время работы с предельными нагрузками, современная наука не располагает, во всяком случае, не спешит поделиться ими с широким кругом читателей. Связано это, на мой взгляд, в первую очередь с тем, что основные исследования всегда были направлены на обеспечение потребностей "классического" спорта, основой которого в большей степени является тренировка выносливости - работоспособности. Максимум, что мне удалось обнаружить, это исследование структурных изменений в мышцах крыс после получасового интенсивного плавания, или бега в колесе. Понятно, что по этим данным тяжело судить о реакции мышц атлета на напряжение, поддерживать которое мышцы способны только несколько секунд. Избавившись от иллюзий найти готовые ответы, я решил самостоятельно путем анализа всех известных мне данных смоделировать процессы, протекающие в мышцах при нагрузках близких к предельным. Разработанная мною модель воздействия тренировки на мышечный аппарат человека оказалась способной описать практически все известные эмпирические факты и позволила впервые найти ответы на ряд вопросов, не получивших до настоящего момента времени сколь ни будь приемлемого объяснения со стороны спортивных физиологов. Например, мне удалось в общих чертах на молекулярном уровне определить механизм возникновения микротравм мышечного волокна (источников посттренировочной боли и общего стрессового воздействия на организм), указать точные условия их возникновения, а так же найти причины адаптации мышц к нагрузке и объяснить феномен снижения посттренировочной боли при регулярных тренировках. Ознакомиться с этой информацией читатель сможет во второй части статьи, в которой, собственно, и изложена суть моей теории, на основе которой в третьей части статьи я постарался объяснить, почему работает "Супертренинг" Ментцера, тренировки в котором ограничиваются лишь одним подходом в упражнении, и для чего предназначены и как работают интервальные (многоподходные) тренировки, в чем секрет лифтерского или штангисткого силового цикла и чем определяется воздействие большинства иных тренировочных методик и приемов. Для неподготовленного читателя прилагаемый материал окажется, скорее всего, слишком сложным для понимания, поэтому, для начала, я рекомендую внимательно изучить первую часть статьи, в которой в доступной форме изложены основные сведения о строении и принципах функционирования мышц, которые мне удалось собрать из специализированных научных трудов советских и российских ученых. Должен, заметить, что те сведения, которые до сего момента времени излагались в популярной литературе (журналах, работах Хетфилда или Тюннеманна и Хартманна), и даже в учебниках по физиологии мышечной деятельности для спортивных ВУЗов, с которыми мне удалось ознакомиться, являются не полными. Как правило, процессы синтеза белка клеткой в данной литературе даже не рассматриваются, в результате многие атлеты, считающие себя специалистами в области "химии", рассуждают о рецепторах стероидных гормонов, об их "забивке", "повышении чувствительности", образовании новых, даже не осознавая, что эффект стероидных гормонов реализуется через воздействие на генетический аппарат, заключенный в ядре клетки, и именно ядро является конечным рецептором тестостерона, кортизола и ряда других гормонов. Не осознают они и то, что увеличить количество рецепторов тестостерона можно только увеличив количество клеточных ядер. Да, ядра мышечных клеток, как и сама мышечная клетка, не способны к делению и размножению, но спортивные физиологи как будто намерено игнорируют информацию о наличии в мышечном волокне клеток - сателлит (несформированных мышечных клеток), сохраняющих способность к делению на протяжении всей жизни человека, благодаря чему и обеспечивается увеличение количества мышечных ядер и регенерация мышечных волокон при механическом повреждении ткани. Как этот факт может повлиять на рост объема и силы мышц читатель так же сможет узнать из второй части.
И так, мне удалось создать более - менее цельную теорию тренинга, на физиологическом уровне объясняющую (конечно в общих чертах) воздействие тренировки на мышечный аппарат человека и позволяющую найти ответы на большинство вопросов, интересующих читателя.
Предвижу сомнения скептиков - человек без специального образования лезет в дебри новой для себя науки, да еще набирается наглости выносить на суд публики собственные теории. Ну что же, если ученым нет дела до проблем бодибилдинга, то приходится полагаться на собственные силы, в конце концов "спасение утопающих - дело рук самих утопающих". И так, если вы готовы, то вперед!
Различают три типа мышечной ткани: скелетная, гладкая и сердечная. Функция сердечной ткани понятна из названия, и ее роль, я думаю, объяснять не надо. О существовании гладких мышц мы зачастую даже не догадываемся, так как это мышцы внутренних органов, и мы лишены возможности напрямую управлять ими, впрочем, как и сердечной мышцей. Между тем, именно гладкие мышцы сокращают стенки сосудов, производят сокращение кишечника, способствуя перемещению пищи, и выполняют множество других жизненно важных функций. Задача скелетных мышц - перемещение частей скелета относительно друг друга (отсюда и название). Именно эти мышцы мы с таким упорством пытаемся нарастить на своем теле, и именно их строение и свойства мы будем рассматривать в дальнейшем.
Как известно, все ткани организма имеют клеточную структуру, не представляют исключение и мышцы. Поэтому мне придется провести краткий экскурс в цитологию - науку о клетке, и напомнить читателям о роли и свойствах основных структур клетки.
В грубом приближении клетка состоит из двух важнейших, взаимосвязанных между собой частей - цитоплазмы и ядра.
Ядро - содержит в себе молекулы ДНК, в которых заключена вся наследственная информация. ДНК - полимер, закрученный в виде двойной спирали, каждая спираль которого составлена из огромного количества четырех видов мономеров, называемых нуклеотидами. Последовательность нуклеотидов в цепочке кодирует все белки организма.
Ядро ответственно за размножение клетки - деление. Деление клетки начинается с разделения молекулы ДНК на две спирали, каждая из которых способна достроить парную из набора свободных нуклеотидов и вновь превратится в молекулу ДНК. Таким образом, количество ДНК в ядре удваивается, далее ядро делится на две части, а за ним и вся клетка.
Цитоплазма - это все, что в клетке окружает ядро. Она состоит из цитозоли (клеточной жидкости), в которую включены различные органеллы, такие как митохондрии, лизосомы, рибосомы и прочие.
Митохондрии - это энергетические станции клетки, в них с помощью различных ферментов происходит окисление углеводов и жирных кислот. Энергия, выделяющаяся при окислении веществ, идет на присоединение третьей фосфатной группы к молекуле Аденезиндифосфата (АДФ) с образованием Аденезинтрифосфата (АТФ) - универсального источника энергии для всех процессов, протекающих в клетке. Отсоединяя третью фосфатную группу и вновь превращаясь в АДФ, АТФ выделяет запасенную ранее энергию.
Ферменты или Энзимы - вещества белковой природы в сотни и тысячи раз увеличивающие скорость протекания химических реакций. Практически все жизненно важные химические процессы в организме происходят только в присутствии специфических ферментов.
Лизосомы - округлые пузырьки, содержащие около 50 ферментов. Лизосомные ферменты расщепляют поглощенный клеткой материал и собственные внутренние структуры клетки (автолизис). Лизосомы, сливаясь в фагосомы, способны переваривать целые органеллы, подлежащие дезинтеграции.
Рибосомы - органеллы, на которых происходит сборка белковой молекулы.
Клеточная мембрана - оболочка клетки, она обладает избирательной проницаемостью, то есть способностью пропускать одни вещества и задерживать другие. Задача мембраны сохранять постоянство внутренней среды клетки.
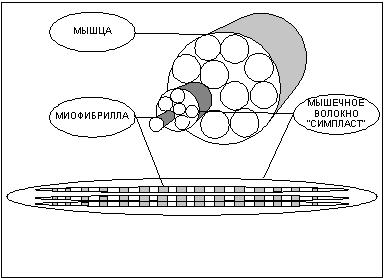
Структурно-функциональной единицей скелетной мышцы является симпласт или мышечное волокно - огромная клетка, имеющая форму протяженного цилиндра с заостренными краями (в дальнейшем под наименованием симпласт, мышечное волокно, мышечная клетка следует понимать один и тот же объект). Длина мышечной клетки чаще всего соответствует длине целой мышцы и достигает 14 см, а диаметр равен нескольким сотым долям миллиметра. Мышечное волокно, как и любая клетка, окружено оболочкой - сарколемой. Снаружи отдельные мышечные волокна окружены рыхлой соединительной тканью, которая содержит кровеносные и лимфатические сосуды, а так же нервные волокна. Группы мышечных волокон образуют пучки, которые, в свою очередь, объединяются в целую мышцу, помещенную в плотный чехол соединительной ткани, переходящей на концах мышцы в сухожилия, крепящиеся к кости.
Усилие, вызываемое сокращением длины мышечного волокна, передается через сухожилия костям скелета и приводит их в движение.
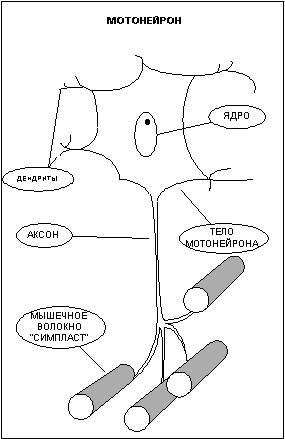
Управление сократительной активностью мышцы осуществляется с помощью большого числа мотонейронов (рис.2) - нервных клеток, тела которых лежат в спинном мозге, а длинные ответвления - аксоны в составе двигательного нерва подходят к мышце. Войдя в мышцу, аксон разветвляется на множество веточек, каждая из которых подведена к отдельному волокну. Таким образом, один мотонейрон иннервирует целую группу волокон (так называемая нейромоторная единица), которая работает как единое целое.
Мышца состоит из множества нервномоторных единиц и способна работать не всей своей массой, а частями, что позволяет регулировать силу и скорость сокращения.
Для понимания механизма сокращения мышцы необходимо рассмотреть внутреннее строение мышечного волокна, которое, как вы уже поняли, сильно отличается от обычной клетки. Начнем с того, что мышечное волокно многоядерно. Связано это с особенностями формирования волокна при развитии плода. Симпласты (мышечные волокна) образуются на этапе эмбрионального развития организма из клеток предшественников - миобластов. Миобласты (неоформленные мышечные клетки) интенсивно делятся, сливаются и образуют мышечные трубочки с центральным расположением ядер. Затем в мышечных трубочках начинается синтез миофибрилл (сократительных структур клетки см. ниже), и завершается формирование волокна миграцией ядер на периферию. Ядра мышечного волокна к этому времени уже теряют способность к делению, и за ними остается только функция генерации информации для синтеза белка.
Но не все миобласты идут по пути слияния, часть из них обособляется в виде клеток-сателлитов, располагающихся на поверхности мышечного волокна, а именно в сарколеме, между плазмолемой и базальной мембраной - составными частями сарколемы. Клетки-сателлиты, в отличие от мышечных волокон, не утрачивают способность к делению на протяжении всей жизни, что обеспечивает увеличение мышечной массы волокон и их обновление. Восстановление мышечных волокон при повреждении мышцы возможно благодаря клеткам-сателлитам. При гибели волокна, скрывающиеся в его оболочке, клетки-сателиты активизируются, делятся и преобразуются в миобласты. Миобласты сливаются друг с другом и образуют новые мышечные волокна, в которых затем начинается сборка миофибрилл. То есть при регенерации полностью повторяются события эмбрионального (внутриутробного) развития мышцы.
Помимо многоядерности отличительной чертой мышечного волокна является наличие в цитоплазме (в мышечном волокне ее принято называть саркоплазмой) тонких волоконец - миофибрилл (рис.1), расположенных вдоль клетки и уложенных параллельно друг другу. Число миофибрилл в волокне достигает двух тысяч. Миофибриллы являются сократительными элементами клетки и обладают способностью уменьшать свою длину при поступлении нервного импульса, стягивая тем самым мышечное волокно. Под микроскопом видно, что миофибрилла имеет поперечную исчерченность - чередующиеся темные и светлые полосы. При сокращении миофибриллы светлые участки уменьшают свою длину и при полном сокращении исчезают вовсе. Для объяснения механизма сокращения миофибриллы около пятидесяти лет назад Хью Хаксли была разработана модель скользящих нитей, затем она нашла подтверждение в экспериментах и сейчас является общепринятой.
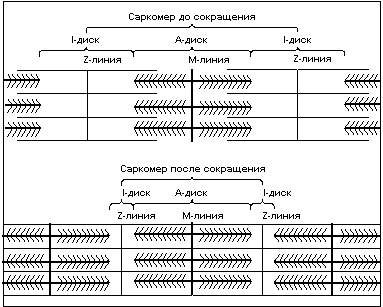
Чередование светлых и темных полос в миофибрильной нити определяется упорядоченным расположением по длине миофибриллы толстых нитей белка миозина и тонких нитей белка актина; толстые нити содержатся только в темных участках (А-диск) (рис.3), светлые участки (I-диск) не содержат толстых нитей, в середине I-диска находится Z-линия к ней крепятся тонкие нити актина. Участок миофибриллы состоящий из А-диска (темной полосы) и двух половинок I-дисков (светлых полос) называют саркомером. Сокращение длины саркомера происходит путем втягивания тонких нитей актина между толстыми нитями миозина. Скольжение нитей актина вдоль нитей миозина происходит благодаря наличию у нитей миозина боковых ответвлений, называемых мостиками. Головка миозинового мостика сцепляется с актином и изменяет угол наклона к оси нити, тем самым как бы продвигая нить миозина и актина относительно друг друга, затем отцепляется, сцепляется вновь и вновь совершает движение. Перемещение миозиновых мостиков можно сравнить с гребками весел на галерах. Как перемещение галеры в воде происходит благодаря движению весел, так и скольжение нитей происходит благодаря гребковым движениям мостиков, существенное отличие состоит лишь в том, что движение мостиков асинхронно.
Тонкая нить представляет из себя две спирально скрученные нити белка актина. В канавках спиральной цепочки залегает двойная цепочка другого белка - тропомиозина. В расслабленном состоянии мостики миозина не имеют возможности связаться с актином, так как места сцепления блокированы тропомиозином. При поступлении по аксону двигательного мотонейрона нервного импульса клеточная мембрана меняет полярность заряда, и из специальных терминальных цистерн, расположенных вокруг каждой миофибриллы вдоль всей ее длины, в саркоплазму выбрасываются ионы кальция (Са++) (рис.4).
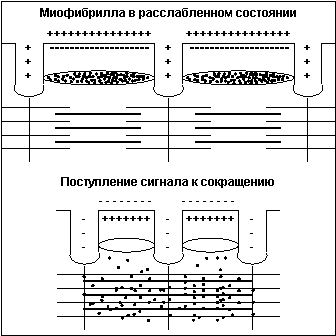
Под воздействием Са++ нить тропомиозина входит глубже в канавку и освобождает места для сцепления миозина с актином, мостики начинают цикл гребков. Сразу после высвобождения Са++ из терминальных цистерн он начинает закачиваться обратно, концентрация Са++ в саркоплазме падает, тропомиозин выдвигается из канавки и блокирует места сцепления мостиков - волокно расслабляется. Новый импульс опять выбрасывает Са++ в саркоплазму и все повторяется. При достаточной частоте импульсации (не менее 20 Гц) отдельные сокращения почти полностью сливаются, то есть достигается состояние устойчивого сокращения, называемое тетаническим сокращением или гладким тетанусом.
Естественно, что для движения мостика требуется энергия. Как я уже упоминал ранее, универсальным источником энергии в живом организме является молекула АТФ. Под действием фермента АТФазы АТФ гидролизуется, отсоединяя фосфатную группу в виде ортофосфорной кислоты (Н3РО4), и превращается в АДФ, при этом высвобождается энергия.
АТФ + H2O = АДФ + H3PO4 + энергия.
Головка миозинового мостика при контакте с актином обладает АТФазной активностью и соответственно возможностью расщеплять АТФ и получать энергию, необходимую для движения.
Запас молекул АТФ в мышце ограничен, поэтому расход энергии при работе мышцы требует постоянного его восполнения. Мышца имеет три источника воспроизводства энергии: расщепление креатинфосфата; гликолиз; окисление органических веществ в митохондриях.
Креатинфосфат обладает способностью отсоединять фосфатную группу и превращаться в креатин, присоединяя фосфатную группу к АДФ, которая превращается в АТФ.
АДФ + креатинфосфат = АТФ + креатин.
Эта реакция получила название - реакции Ломана. Запасы креатинфосфата в волокне не велики, поэтому он используется в качестве источника энергии только на начальном этапе работы мышцы, до момента активизации других более мощных источников - гликолиза и кислородного окисления. По окончании работы мышцы реакция Ломана идет в обратном направлении, и запасы креатинфосфата в течение нескольких минут восстанавливаются.
Гликолиз - процесс распада одной молекулы глюкозы (C6H12O6) на две молекулы молочной кислоты (C3H6O3) с выделением энергии, достаточной для "зарядки" двух молекул АТФ, протекает в саркоплазме под воздействием 10 специальных ферментов.
C6H12O6 + 2H3PO4 + 2АДФ = 2C3H6O3 + 2АТФ + 2H2O.
Гликолиз протекает без потребления кислорода (такие процессы называются анаэробными) и способен быстро восстанавливать запасы АТФ в мышце.
Окисление протекает в митохондриях под воздействием специальных ферментов и требует затрат кислорода, а соответственно и времени на его доставку. Такие процессы называются аэробными. Окисление происходит в несколько этапов, сначала идет гликолиз (см. выше), но образовавшиеся в ходе промежуточного этапа этой реакции две молекулы пирувата не преобразуются в молекулы молочной кислоты, а проникают в митохондрии, где окисляются в цикле Кребса до углекислого газа СО2 и воды Н2О и дают энергию для производства еще 36 молекул АТФ. Суммарное уравнение реакции окисления глюкозы выглядит так:
C6H12O6 + 6O2 + 38АДФ + 38H3PO4 = 6CO2 + 44H(2)О + 38АТФ.
Итого распад глюкозы по аэробному пути дает энергию для восстановления 38 молекул АТФ. То есть окисление в 19 раз эффективнее гликолиза.
Скелетные мышцы и образующие их мышечные волокна различаются по множеству параметров: скорости сокращения, утомляемости, диаметру, цвету и т.д. Традиционно выделяют красные и белые, медленные и быстрые, гликолитические и окислительные волокна.
Скорость сокращения мышечного волокна определяются типом миозина. Изоформа миозина, обеспечивающая высокую скорость сокращения, - быстрый миозин характеризуется высокой активностью АТФазы, а соответственно и скоростью расхода АТФ. Изоформа миозина с меньшей скоростью сокращения - медленный миозин, характеризуется меньшей активностью АТФазы. Волокна, с высокой активностью АТФазы и скоростью расхода АТФ, принято называть быстрыми волокнами, волокна, характеризующиеся низкой активностью АТФазы и меньшей скоростью расхода АТФ, - медленными волокнами.
Для восполнения затрат энергией мышечные волокна используют окислительный либо гликолитический путь образования АТФ.
Окислительные, или красные, мышечные волокна небольшого диаметра окружены массой капилляров и содержат много белка миоглобина (именно наличие этого белка придает волокнам красный цвет). Многочисленные митохондрии красных волокон имеют высокий уровень активности окислительных ферментов. Мощная сеть капилляров необходима для доставки с кровью большого количества кислорода, а миоглобин используется для транспортировки кислорода внутри волокна от поверхности к митохондриям. Энергию красные волокна получают путем окисления в митохондриях углеводов и жирных кислот.
Гликолитические, или белые, мышечные волокна имеют больший диаметр, в их саркоплазме содержится значительное количество гранул гликогена, митохондрии не многочисленны, активность окислительных ферментов значительно уступает активности гликолитических. Гликоген, его еще принято называть "животным крахмалом", - сложный полисахорид с высокой молекулярной массой служит резервным питательным веществом белого волокна. Гликоген распадается до глюкозы, которая, служит топливом при гликолизе.
Быстрые волокна, обладающие высокой активностью АТФазы и соответственно скоростью расхода энергии, требуют высокой скорости воспроизводства АТФ, обеспечить которую может только гликолиз, так как, в отличие от окисления, он протекает непосредственно в саркоплазме и не требует времени на доставку кислорода к митохондриям и доставку энергии от них к миофибриллам. Поэтому быстрые волокна предпочитают гликолитический путь воспроизводства АТФ и соответственно относятся к белым волокнам. За высокую скорость получения энергии белые волокна платят быстрой утомляемостью, так как гликолиз, как видно из уравнения реакции ведет к образованию молочной кислоты, накопление которой повышает кислотность среды и вызывает усталость мышцы и в конечном итоге останавливает ее работу.
Медленные волокна, характеризующиеся низкой активностью АТФазы, не требуют столь быстрого восполнения запасов АТФ и для обеспечения потребности в энергии используют путь окисления, то есть относятся к красным волокнам. Благодаря этому медленные волокна являются низко утомляемыми и способны поддерживать относительно небольшое, но длительное напряжение.
Существует и промежуточный тип волокон с высокой АТФазной активностью, и окислительно-гликолитическим путем воспроизводства АТФ.
Тип мышечного волокна зависит от мотонейрона его иннервирующего. Все волокна одного мотонейрона принадлежат к одному типу. Интересный факт - при подключении к быстрому волокну аксона медленного мотонейрона и наоборот, волокно перерождается, меняя свой тип. До недавнего времени существовало две точки зрения на зависимость типа волокна от типа мотонейрона, одни исследователи полагали, что свойства волокна определяются частотой импульсации мотонейрона, другие, что эффект влияния на тип волокна определяется гормоноподобными веществами поступающими из нерва (эти вещества до настоящего времени не выделены). Исследования последних лет показывают, что обе точки зрения имеют право на существование, воздействие мотонейрона, на волокно осуществляется обоими способами.
Как вы знаете на собственном опыте, человек обладает возможностью произвольно регулировать силу и скорость мышечного сокращения. Реализуется эта возможность несколькими способами. С одним из них вы уже знакомы - это управление частотой нервной импульсации. Подавая волокну одиночные команды на сокращение, можно добиться в нем легкого напряжения. Так, например, поддерживающие позу мышцы слегка напряжены, даже когда человек отдыхает. Повышая частоту импульсов можно увеличивать силу сокращения до максимально возможной для данного волокна в данных условиях работы, которая достигается при слиянии отдельных импульсов в тетанус.
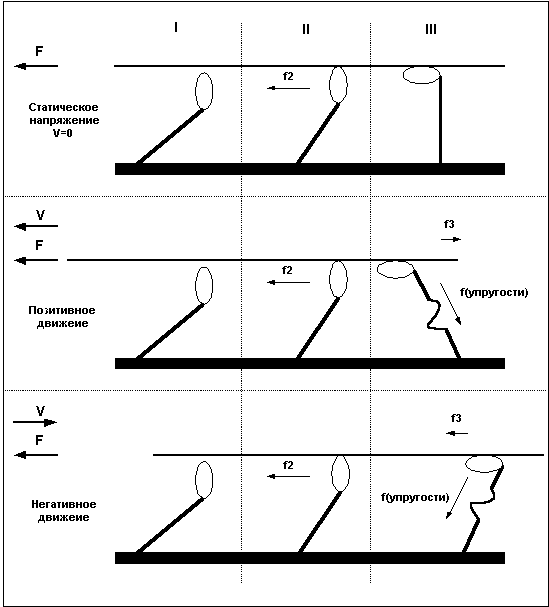
Сила, развиваемая волокном в состоянии тетануса, не всегда одинакова и зависит от характера и скорости движения. При статическом напряжении (когда длина волокна остается постоянной) сила, развиваемая волокном, больше чем при сокращении волокна, и чем быстрее сокращается волокно, тем меньшую силу оно способно развить. Максимальную же силу волокно развивает в условиях негативного движения, то есть при удлинении волокна.
При отсутствии внешней нагрузки волокно сокращается с максимальной скоростью. При увеличении нагрузки скорость сокращения волокна уменьшается и по достижении определенного уровня нагрузки падает до нуля, при дальнейшем увеличении нагрузки волокно удлиняется.
Причину в различии силы волокна при различных направлениях движения легко понять, рассмотрев уже приведенный ранее пример с гребцами и веслами. Дело в том, что после завершения "гребка" миозиновый мостик некоторое время находится в состоянии сцепления с нитью актина, представьте, что весло после гребка так же не сразу извлекается из воды, а находится погруженным еще некоторое время. В случае, когда гребцы плывут вперед (позитивное движение), весла, остающиеся погруженными в воду после завершения гребка, тормозят движение и мешают плыть, в то же время, если лодка буксируется назад, а гребцы сопротивляются этому движению, то погруженные весла так же мешают движению, и буксиру приходится прилагать большие усилия. То есть, при сокращении волокна, сцепленные мостики мешают движению и ослабляют силу волокна, при негативном движении - удлинении мышцы - не отцепленные мостики так же мешают движению, но в этом случае они как бы поддерживают опускающийся вес, что и позволяет волокну развивать большую силу. Легче всего понять различия между статическим напряжением, позитивным и негативным движением взглянув на Рис.5.
Итак, мы рассмотрели основные факторы, влияющие на силу и скорость сокращения отдельного волокна. Сила же сокращения целой мышцы зависит от количества волокон в данный момент времени вовлеченных в работу.
При поступлении от ЦНС (центральной нервной системы) к мотонейронам (расположенным в спинном мозге), возбуждающего сигнала (запускающий импульс), мембрана мотонейрона поляризуется и он генерирует серию импульсов, направляемых по аксону к волокнам. Чем сильнее воздействие на мотонейрон (поляризация мембраны), тем выше частота генерируемого им импульса - от небольшой стартовой частоты (4-5 Гц), до максимально возможной, для данного мотонейрона, частоты (50 Гц и более). Быстрые мотонейроны способны генерировать гораздо более высокочастотный импульс, чем медленные, поэтому сила сокращения быстрых волокон гораздо больше подвержена частотной регуляции, чем сила медленных.
В то же время имеется и обратная связь с мышцей, от которой поступают тормозящие сигналы, уменьшающие поляризацию мембраны мотонейрона и снижающие его ответ.
Каждый мотонейрон имеет свой порог возбудимости. Если сумма возбуждающих и тормозящих сигналов, превышает этот порог и на мембране достигается необходимый уровень поляризации, то мотонейрон вовлекается в работу. Медленные мотонейроны имеют, как правило, низкий порог возбудимости, а быстрые высокий. Мотонейроны же целой мышцы имеют широкий спектр значений этого параметра. Таким образом, при повышении силы сигнала ЦНС, активируется все большее число мотонейронов, а мотонейроны с низким порогом возбудимости увеличивают частоту генерируемого импульса.
Когда требуется легкое усилие, например, при ходьбе или беге трусцой, активируется небольшое число медленных мотонейронов и соответствующее число медленных волокон, ввиду высокой выносливости этих волокон такая работа может поддерживаться очень долго. По мере увеличения нагрузки ЦНС приходится посылать все более сильный сигнал, и большее число мотонейронов (а, следовательно, волокон), вовлекается в работу, а те, что уже работали, увеличивают силу сокращения, по причине увеличения частоты импульсации, поступающей от мотонейронов. По мере увеличения нагрузки в работу включаются быстрые окислительные волокна, а по достижения определенного уровня нагрузки (20%-25% от максимума), например, во время подъема в гору или финального спурта, силы окислительных волокон становится недостаточно, и посылаемый ЦНС сигнал включает в работу быстрые - гликолитические волокна. Быстрые волокна значительно повышают силу сокращения мышцы, но, в свою очередь, быстро утомляются, и в работу вовлекается все большее их количество. Если уровень внешней нагрузки не уменьшается, работу в скором времени приходится останавливать из за усталости, в результате накопления молочной кислоты.
Скорость движения мышцы зависит от соотношения внешней нагрузки и количества вовлеченных в движение волокон. Мобилизация относительно небольшого количества волокон, приведет к медленному движению, как только совместная сила сокращающихся волокон превысит уровень внешней силы. Мобилизация все большего количества волокон при той же внешней нагрузке приведет к увеличению скорости сокращения, а увеличение внешней нагрузки, или падение силы волокон в результате усталости, при неизменном количестве мобилизованных волокон, приведет к падению скорости сокращения.
При предельных нагрузках, например, при подъеме максимального веса или подъеме относительно небольшого веса, но с максимальной скоростью, сокращается сразу максимально возможное для данного индивида число волокон.
По вопросу вовлечения волокон при предельных нагрузках в литературе приводятся противоречивые сведения. Так ряд авторов утверждает, что даже у тренированных спортсменов при предельных нагрузках активно не более 85% от общего количества двигательных единиц в мышце. С другой стороны, Гурфинкель и Левин приводят данные, что большая часть двигательных единиц рекрутируется уже при нагрузках до 50% от максимума, и в дальнейшем, при повышении силы, подключается лишь небольшая часть (около 10%) самых крупных двигательных единиц, а рост силы от 75% до 100% происходит уже не за счет вовлечения новых двигательных единиц, а за счет повышения частоты импульса, генерируемого мотонейронами. Независимо от того, чья точка зрения является правильной, важно одно - сила мышцы зависит от силы импульса ЦНС, а чем обусловлен рост силы при нагрузках свыше 75% от максимальной только ли ростом частоты, генерируемой мотонейронами, или еще и вовлечением в работу новых мотонейронов не принципиально. Важно так же и то, что в мышцу встроен ограничительный механизм контроля за развиваемым напряжением, осуществляется через сухожильные органы Гольджи. Сухожильные рецепторы регистрируют напряжение и при превышении критического значения оказывают тормозящее воздействие на мотонейроны. Считается, что при отключении контроля за напряжением проявляется "сверхсила" человека, регистрируемая в экстремальных ситуациях.
|
| Реклама&Меню | © 1999-2026 ATHLETE.RU |